Wenn Sie das Fenster schließen, wird Ihre Konfiguration nicht gespeichert, es sei denn, Sie haben Ihren Artikel in die Bestellung aufgenommen oder zu Ihren Favoriten hinzugefügt.
Klicken Sie auf OK, um das MILLIPLEX® MAP-Tool zu schließen oder auf Abbrechen, um zu Ihrer Auswahl zurückzukehren.
Wählen Sie konfigurierbare Panels & Premixed-Kits - ODER - Kits für die zelluläre Signaltransduktion & MAPmates™
Konfigurieren Sie Ihre MILLIPLEX® MAP-Kits und lassen sich den Preis anzeigen.
Konfigurierbare Panels & Premixed-Kits
Unser breites Angebot enthält Multiplex-Panels, für die Sie die Analyten auswählen können, die am besten für Ihre Anwendung geeignet sind. Unter einem separaten Register können Sie das Premixed-Cytokin-Format oder ein Singleplex-Kit wählen.
Kits für die zelluläre Signaltransduktion & MAPmates™
Wählen Sie gebrauchsfertige Kits zur Erforschung gesamter Signalwege oder Prozesse. Oder konfigurieren Sie Ihre eigenen Kits mit Singleplex MAPmates™.
Die folgenden MAPmates™ sollten nicht zusammen analysiert werden: -MAPmates™, die einen unterschiedlichen Assaypuffer erfordern. -Phosphospezifische und MAPmate™ Gesamtkombinationen wie Gesamt-GSK3β und Gesamt-GSK3β (Ser 9). -PanTyr und locusspezifische MAPmates™, z.B. Phospho-EGF-Rezeptor und Phospho-STAT1 (Tyr701). -Mehr als 1 Phospho-MAPmate™ für ein einziges Target (Akt, STAT3). -GAPDH und β-Tubulin können nicht mit Kits oder MAPmates™, die panTyr enthalten, analysiert werden.
.
Bestellnummer
Bestellinformationen
St./Pkg.
Liste
Dieser Artikel wurde zu Ihren Favoriten hinzugefügt.
Wählen Sie bitte Spezies, Panelart, Kit oder Probenart
Um Ihr MILLIPLEX® MAP-Kit zu konfigurieren, wählen Sie zunächst eine Spezies, eine Panelart und/oder ein Kit.
Custom Premix Selecting "Custom Premix" option means that all of the beads you have chosen will be premixed in manufacturing before the kit is sent to you.
Catalogue Number
Ordering Description
Qty/Pack
List
Dieser Artikel wurde zu Ihren Favoriten hinzugefügt.
Spezies
Panelart
Gewähltes Kit
Menge
Bestellnummer
Bestellinformationen
St./Pkg.
Listenpreis
96-Well Plate
Menge
Bestellnummer
Bestellinformationen
St./Pkg.
Listenpreis
Weitere Reagenzien hinzufügen (MAPmates erfordern die Verwendung eines Puffer- und Detektionskits)
Menge
Bestellnummer
Bestellinformationen
St./Pkg.
Listenpreis
48-602MAG
Buffer Detection Kit for Magnetic Beads
1 Kit
Platzsparende Option Kunden, die mehrere Kits kaufen, können ihre Multiplex-Assaykomponenten in Kunststoffbeuteln anstelle von Packungen erhalten, um eine kompaktere Lagerung zu ermöglichen.
Dieser Artikel wurde zu Ihren Favoriten hinzugefügt.
Das Produkt wurde in Ihre Bestellung aufgenommen
Sie können nun ein weiteres Kit konfigurieren, ein Premixed-Kit wählen, zur Kasse gehen oder das Bestell-Tool schließen.
This Anti-IRF3 Antibody, clone SL-12 is validated for use in Western Blotting and Immunocytochemistry and Immunoprecipitation for the detection of IRF3.
More>>This Anti-IRF3 Antibody, clone SL-12 is validated for use in Western Blotting and Immunocytochemistry and Immunoprecipitation for the detection of IRF3. Less<<
Anti-IRF3 Antibody, clone SL-12: SDB (Sicherheitsdatenblätter), Analysenzertifikate und Qualitätszertifikate, Dossiers, Broschüren und andere verfügbare Dokumente.
Interferon regulatory factors (IRFs) comprise a family of transcription factors that function with the Jak/Stat pathway to regulate interferon (IFN) and IFN-inducible gene expression in response to viral infection. IRF-3 can inhibit cell growth and plays a critical role in controlling the expression of genes in the innate immune response. In unstimulated cells, IRF-3 is present in the cytoplasm. Viral infection results in phosphorylation of IRF-3 and leads to its translocation to the nucleus where it activates promoters containing IRF-3-binding sites. Phosphorylation of IRF-3 occurs at a cluster of C-terminal serine and threonine residues (between 385 and 405) leading to its association with the p300/CBP coactivator protein that promotes DNA binding and transcriptional activity.
References
Product Information
Format
Purified
Presentation
Purified mouse monoclonal IgG1κ in buffer containing 0.1 M Tris-Glycine (pH 7.4), 150 mM NaCl with 0.05% sodium azide.
This Anti-IRF3 Antibody, clone SL-12 is validated for use in Western Blotting and Immunocytochemistry and Immunoprecipitation for the detection of IRF3.
Key Applications
Western Blotting
Immunocytochemistry
Immunoprecipitation
Application Notes
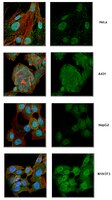
Immunocytochemistry Analysis: A 1:50 dilution from a representative lot detected IRF3 in HeLa, A-431, HepG2, and NIH/3T3 cells. DyLight 488® is a registered trademark of Thermo Fisher Scientific. Immunoprecipitation Analysis: A representative lot immunoprecipitated IRF3 in HEC1B cells treated or untreated with virus and M059J DNA-PK - and M059J cells with DNA-PK+ (Karpova, A.Y., et al. (2002). PNAS. 99(5):2818-2823). Western Blotting Analysis: A representative lot detected IRF3 in HFK (human foreskin keratinocytes) cells expressing BPV ORF's (Ronco, L.V., et al. (1998) Genes Dev. 12:2061-2072). Western Blotting Analysis: A representative lot detected IRF3 in Transfected Mock and SeV infected 293 cells (Basler, C.F., et al. (2003) Journal of Virology. 77(14):7945-7956). Immunocytochemistry Analysis: A representative lot detected IRF3 in hec-1b cells infected with delNS1 or PR8 (Talon. J., et al. (2000). Journal of Virology. 74(17):7989-7996). Immunocytochemistry Analysis: A representative lot detected IRF3 in HEC1B cells treated or untreated with virus (Karpova, A.Y., et al. (2002). PNAS. 99(5):2818-2823).
~49 kDa observed. IRF3 has isoforms at ~49 kDa, ~47 kDa, ~33 kDa, ~16 kDa, & ~12 kDa.
Physicochemical Information
Dimensions
Materials Information
Toxicological Information
Safety Information according to GHS
Safety Information
Product Usage Statements
Quality Assurance
Evaluated by Western Blotting in HeLa cell lysate.
Western Blotting Analysis: 1.0 µg/mL of this antibody detected IRF3 in 10 µg of HeLa cell lysate.
Usage Statement
Unless otherwise stated in our catalog or other company documentation accompanying the product(s), our products are intended for research use only and are not to be used for any other purpose, which includes but is not limited to, unauthorized commercial uses, in vitro diagnostic uses, ex vivo or in vivo therapeutic uses or any type of consumption or application to humans or animals.
The Ebola virus VP35 protein was previously found to act as an interferon (IFN) antagonist which could complement growth of influenza delNS1 virus, a mutant influenza virus lacking the influenza virus IFN antagonist protein, NS1. The Ebola virus VP35 could also prevent the virus- or double-stranded RNA-mediated transcriptional activation of both the beta IFN (IFN-beta) promoter and the IFN-stimulated ISG54 promoter (C. Basler et al., Proc. Natl. Acad. Sci. USA 97:12289-12294, 2000). We now show that VP35 inhibits virus infection-induced transcriptional activation of IFN regulatory factor 3 (IRF-3)-responsive mammalian promoters and that VP35 does not block signaling from the IFN-alpha/beta receptor. The ability of VP35 to inhibit this virus-induced transcription correlates with its ability to block activation of IRF-3, a cellular transcription factor of central importance in initiating the host cell IFN response. We demonstrate that VP35 blocks the Sendai virus-induced activation of two promoters which can be directly activated by IRF-3, namely, the ISG54 promoter and the ISG56 promoter. Further, expression of VP35 prevents the IRF-3-dependent activation of the IFN-alpha4 promoter in response to viral infection. The inhibition of IRF-3 appears to occur through an inhibition of IRF-3 phosphorylation. VP35 blocks virus-induced IRF-3 phosphorylation and subsequent IRF-3 dimerization and nuclear translocation. Consistent with these observations, Ebola virus infection of Vero cells activated neither transcription from the ISG54 promoter nor nuclear accumulation of IRF-3. These data suggest that in Ebola virus-infected cells, VP35 inhibits the induction of antiviral genes, including the IFN-beta gene, by blocking IRF-3 activation.
Eukaryotic cells have evolved complex signaling networks to sense environmental stress and to repair stress-induced damage. IFN regulatory factor-3 (IRF-3) is a transcription factor that plays a central role in the host response to viral infection. Although the main activity of IRF-3 characterized to date has been its role in the induction of IFN-alpha and -beta after virus infection, recent evidence indicates additional roles for IRF-3 in the response to DNA damage and in virus-induced apoptosis. Here we identify IRF-3 as the first in vivo target for DNA-dependent protein kinase (DNA-PK). Phosphorylation of IRF-3 by DNA-PK after virus infection results in its nuclear retention and delayed proteolysis. These results expand the known roles of DNA-PK and provide a functional link between the cellular machineries that regulate the innate immune response and that sense and respond to DNA damage. As such this study contributes to a more integrated view of the cellular responses to various cellular stress signals.
We present a novel mechanism by which viruses may inhibit the alpha/beta interferon (IFN-alpha/beta) cascade. The double-stranded RNA (dsRNA) binding protein NS1 of influenza virus is shown to prevent the potent antiviral interferon response by inhibiting the activation of interferon regulatory factor 3 (IRF-3), a key regulator of IFN-alpha/beta gene expression. IRF-3 activation and, as a consequence, IFN-beta mRNA induction are inhibited in wild-type (PR8) influenza virus-infected cells but not in cells infected with an isogenic virus lacking the NS1 gene (delNS1 virus). Furthermore, NS1 is shown to be a general inhibitor of the interferon signaling pathway. Inhibition of IRF-3 activation can be achieved by the expression of wild-type NS1 in trans, not only in delNS1 virus-infected cells but also in cells infected with a heterologous RNA virus (Newcastle disease virus). We propose that inhibition of IRF-3 activation by a dsRNA binding protein significantly contributes to the virulence of influenza A viruses and possibly to that of other viruses.
Human papillomavirus 16 E6 oncoprotein binds to interferon regulatory factor-3 and inhibits its transcriptional activity. Ronco, L V, et al. Genes Dev., 12: 2061-72 (1998)
1998
Interferon regulatory factor-3 (IRF-3) was found to specifically interact with HPV16 E6 in a yeast two-hybrid screen. IRF-3 is activated by the presence of double-stranded RNA or by virus infection to form a stable complex with other transcriptional regulators that bind to the regulatory elements of the IFNbeta promoter. We show that IRF-3 is a potent transcriptional activator and demonstrate that HPV16 E6 can inhibit its transactivation function. The expression of HPV16 E6 in primary human keratinocytes inhibits the induction of IFNbeta mRNA following Sendai virus infection. The binding of HPV16 E6 to IRF-3 does not result in its ubiquitination or degradation. We propose that the interaction of E6 with IRF-3 and the inhibition of IRF-3's transcriptional activity may provide the virus a means to circumvent the normal antiviral response of an HPV16-infected cell.